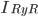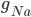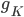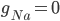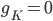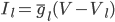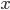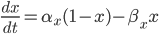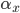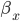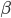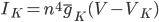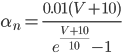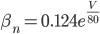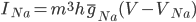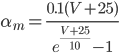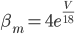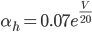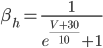I, like many other people, have a passionate and deeply loving relationship with notebooks. All that notebooks and I ask is that you respect our love for the sacred union it represents. There have been a spate of birthdays amongst my friends recently, so I put together some hand-bound notebooks for them.

This one is bound using coptic stitch in gold thread. The cover is made from a repurposed purple pinstriped suit jacket.

I bought the corner protectors from this eBay shop.

The covers are a couple of centimetres wider and longer than the pages.

The signatures are encased in purple, blue, and grey cover papers.

I used this super happy kawaii wrapping paper (from Primark) to line the insides of the covers.

I made it for my awesome friend Sacha, who is a composer. The pages are a mixture of manuscript paper, squared paper, plain, and coloured paper.

I made a PDF of the manuscript paper in LibreOffice Draw. You can download it from here.

The next three notebooks were for my chums Emma, Benji, and Laura. The red fabric used to be a dress, and the blue was a pillowcase.

Manuscript paper for Laura, because she's a classical guitarist, folk musician and composer.

Lined paper for Emma because she's a writer (PDF).

And storyboards for Benji because he's a filmmaker (PDF).

SUPER COOL COVER PAPERS (also from Primark).

I used a mixture of coptic stitch and french tape binding on these books. I learnt the exposed tape technique from this amazing tutorial.

The covers on these books are the same size as the inner papers. I think I prefer making them this way since it's a more economical use of my A4 pieces of cardboard (cannibalised from notebooks and A4 refill pads).



I really like the look of the exposed tape bindings, but I think I prefer to attach covers using coptic stitch as it's a lot less fiddly.

 The EAD begins at around 0.3 seconds.
The EAD begins at around 0.3 seconds. The DAD begins at around 2 seconds. The sarcoplasmic reticulum calcium release current is labelled as
The DAD begins at around 2 seconds. The sarcoplasmic reticulum calcium release current is labelled as